Globicephala macrorhynchus sibo. Tipo Shiho, que se distribuiría en el Océano Pacífico este y norte de Japón.
Globicephala macrorhynchus macrorhynchus. Tipo Naisa, que se distribuiría en el resto de su área de distribución, encontrándose diferencias entre la población del Atlántico y la población del Pacífico e Indico (Van Cise et al. 2019).
Foto: As. Tonina
Taxonomía
El género Globicephala (del Latín ‘globus’ = globo y el Griego ‘kephale’ = cabeza) incluye dos especies desde el año 1971, cuando van Bree demostró que solo existían dos morfotipos reconocibles por diferencias osteológicas:
Calderón tropical o de aleta corta (G. macrorhynchus, Gray 1846)
Calderón común o de aleta larga (G. melaena, Traill 1809).
Este género ha sido revisado en varias ocasiones. Así por ejemplo, Jones et al. (1986) proponen la revisión del nombre específico del calderón común y en la actualidad el nombre específico aceptado es G. melas. Algunas variaciones en la morfología del calderón tropical llevaron a algunos autores a sugerir la existencia de otras especies como G. scammonii (revisado en Bernard & Reilly, 1999), que en la actualidad se considera una variación geográfica del calderón tropical.
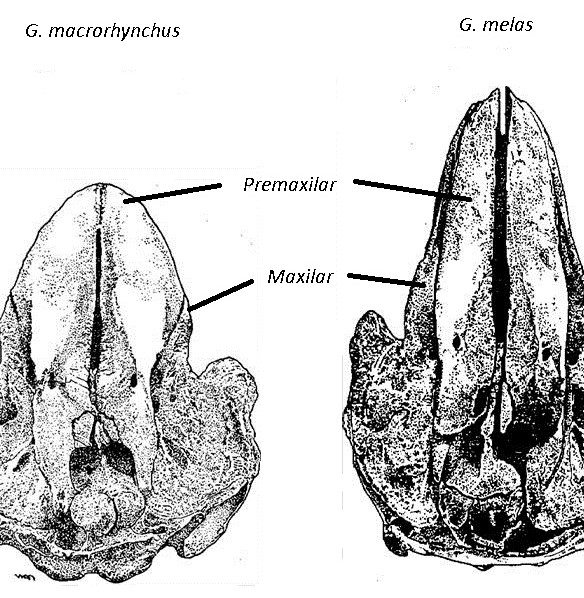
Las dos especies de calderón muestran una apariencia externa similar, diferenciándose principalmente en: la longitud relativa de las aletas pectorales con respecto al cuerpo (más del 16% de la longitud total en G. melas y menos del 15% en G. macrorhynchus), y el número de dientes (9-12 en G.melas y 7-9 en G. macrorhynchus) (Fraser, 1950; Sergeant, 1962; Van Bree, 1971). También se observan diferencias en la aleta dorsal de los machos adultos, que es más alta en los calderones tropicales. Sin embargo, existe solapamiento en estos caracteres (Bloch et al., 1993) y las diferencias en la forma del cráneo son la característica más fiable (Bernard & Reilly, 1999) (Fig.1).
Figura 1. Diferencias en el cráneo de G. macrorhynchus y G. melas.
Un estudio utilizando marcadores genéticos mitocondriales y nucleares ha descubierto que puede darse hibridación entre las dos especies (Miralles et al., 2013), confirmando la existencia de un individuo híbrido adulto, fruto del cruce entre un calderón común y un individuo híbrido fértil, cuya madre era un calderón común y el padre uno tropical. Esta es la primera vez que se describe genéticamente la existencia de un híbrido viable entre estas dos especies en condiciones naturales (Miralles et al., 2013), aunque no es la primera vez que se da el caso entre especies de cetáceos en cautividad (Zornetzer & Duffield, 2003).
Descripción morfológica
El calderón tropical también es llamado ballena piloto, por su traducción directa del nombre inglés “pilot whale”, y “roas” o “roas negro” en Canarias. Es una especie de cuerpo largo y robusto, sin el hocico pronunciado característico de otras especies de delfínidos. Tiene la boca ínfera y con la frente entre bulbosa y cuadrada, dependiendo del género, la edad y la subespecie.
Su patrón de coloración es muy característico y dependiente de la edad.
Los individuos jóvenes son de coloración parduzca o gris claro, que se vuelve negro azabache o gris oscuro con el paso de los años (Yonekura et al., 1980). Presentan áreas claras en la parte ventral del cuerpo, donde poseen una mancha en forma de ancla que empieza bajo la garganta y acaba detrás del ano (Mitchell, 1970). En las hembras esta mancha ventral se extiende alrededor de las hendiduras mamarias (Yonekura et al., 1980) y se ha especulado que podría servir para guiar a las crías durante la lactancia (Mitchell, 1970). Detrás de la aleta dorsal presentan además una mancha gris clara en forma de silla de montar, y muchos individuos poseen además líneas claras post-orbitales (Mitchell, 1970; Kasuya et al., 1988).
La aleta dorsal está situada en la mitad anterior del cuerpo y su forma es muy característica, siendo en general de base ancha y con forma de hoz. Las aletas pectorales, situadas posteriores a la cabeza, son largas, terminadas en punta y con un borde curvo a modo de “codo” (Sergeant, 1962). El pedúnculo caudal es grueso, acabado en una ancha aleta caudal con una muesca separando los dos lóbulos, de bordes cóncavos y con los extremos apuntados (Sergeant, 1962).
Esta especie muestra un marcado dimorfismo sexual.
Los machos adultos alcanzan tamaños que varían entre los 7,2 m en el norte de Japón a los 5,25 m en el sur de Japón, mientras que las hembras alcanzan tamaños de 5,3 a 4,1 m en las mismas zonas geográficas (Kasuya et al., 1988). Las muestras analizadas de calderones tropicales de Tenerife, indican que los calderones de Canarias pertenecen a la subespecie Globicephala macrorhynchus macrorhynchus (Tipo Naisa) (Van Cise et al. 2019).
Figura 2. Diferencias en la cabeza entre un macho y una hembra de G. macrorhynchus.
Las diferencias morfológicas que presenta esta especie y observaciones sobre su comportamiento llevadas a cabo en el medio natural, permiten diferenciar cinco clases de edad/sexo (revisado en Heimlich-Boran, 1993): macho adulto, hembra adulta, adulto indeterminado, juvenil y cría o neonato.
Foto: Bonadea II
Distribución y migraciones
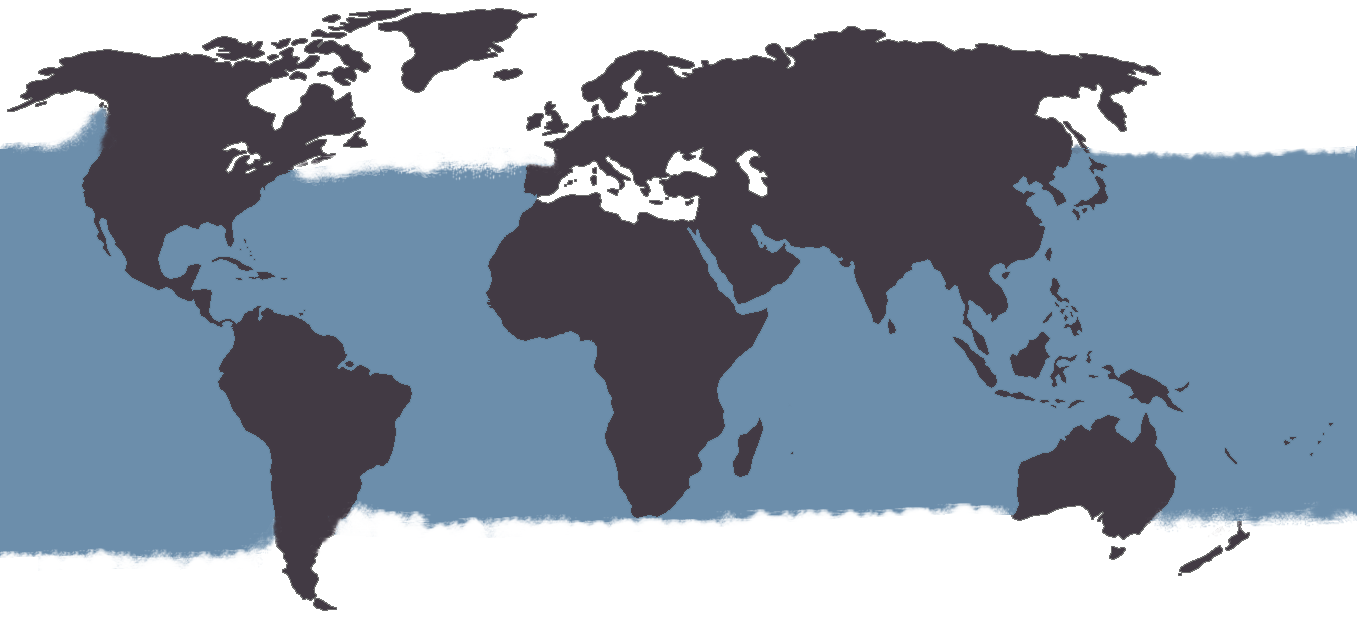
El calderón tropical es una especie oceánica, siendo frecuente observarlo asociado al borde de la plataforma continental, aunque en ocasiones puede acercarse mucho a la costa. Tiene una distribución pantropical en las aguas tropicales y templado cálidas de todos los océanos, aunque puede llegar a latitudes altas, extendiéndose desde aproximadamente los 41º S hasta los 45º N (Bernard & Reilly, 1999) (Fig. 3). Esto provoca que exista cierto solapamiento en la distribución de las dos especies de calderón, lo que, unido a la similitud morfológica de ambas especies, dificulta la determinación precisa de su rango de distribución (Revisado en Bernard & Reilly, 1999).
Figura 3. Distribución del calderón tropical (Globicephala macrorhynchus).
En el Pacífico norte, donde no se encuentran calderones comunes, el rango de distribución del calderón tropical se extiende hacia aguas frías tan al norte como puede ser Hokkaido a 50ºN o la península de Alaska a 57ºN. Se ha sugerido que los calderones tropicales están en proceso de expansión hacia el norte del Pacífico para ocupar los nichos vacíos dejados por los calderones comunes (Bernard & Reilly, 1999). El límite de distribución por el Sur del Pacífico Oeste parece encontrarse a la altura del Oeste de Australia, mientras que por el Este su límite se encontraría en Méjico (revisado en Bernard & Reilly, 1999).
En el Atlántico, el calderón tropical se extiende desde las costas de Sao Paulo en Brasil hasta las americanas de New Jersey por el Oeste y desde Cape Province en Sudáfrica hasta Charente-Maritime en Francia por el Este (Heimlich-Boran, 1993; Bernard & Reilly, 1999).
Esta especie no parece entrar en el Mediterráneo, donde existen poblaciones de calderón comunes que se observan con regularidad, incluyendo zonas de aguas españolas en la región de Murcia, el Mar de Alborán y el Estrecho de Gibraltar (Cañadas, 2006; De Stephanis et al., 2008; Notarbartolo di Sciara & Birkun, 2010).
Apenas existen datos acerca de la distribución del calderón tropical en el Océano Índico, existiendo unos pocos informes sobre avistamientos y varamientos aislados (revisado en Heimlich-Boran, 1993).
Abundancia
No existe una estima para la población mundial de calderón tropical, aunque se considera globalmente abundante (Reeves, 2003). La mayoría de las estimas de abundancia existentes para esta especie, derivan de trabajos realizados en zonas concretas cerca de la costa.
Utilizando la metodología de transectos lineales, se estimó la población de calderón tropical en aguas tropicales del Pacífico Este en 160.200 (Wade & Gerrodette, 1993). En Japón se obtuvieron estimas de 5.300 para la población del norte y 53.000 para la del sur (Miyashita et al., 1996). Se estiman 7.700 individuos en el este del mar de Sulu (Filipinas) (Dolar et al., 2006) y en el Pacífico tropical oriental, la estima más reciente realizada durante el año 2.000, es de 589.000 individuos (Gerrodette & Forcada, 2002). En la costa oeste de Estados Unidos, la abundancia de calderón tropical es variable y parece estar influenciada por las condiciones oceanográficas predominantes (Forney, 1997; Forney & Barlow, 1998, Barlow, 2016). La última estima realizada en la zona, utilizando la media geométrica de las estimaciones de 2008 y estudios de transectos lineares llevados a cabo durante 2014, es de 236 (CV=0,79) animales (Barlow, 2016).
Los datos de fotoidentificación, telemetría, acústicos y genéticos, sugieren que en Hawái residen al menos dos poblaciones de calderón tropical, demográficamente independientes. Los reavistamientos y análisis de redes sociales de individuos fotografiados frente a la isla de Hawái, sugieren la existencia de un gran cluster social y varios más pequeños, con algunos individuos de los cluster pequeños que son comúnmente reavistados en las aguas de Hawái (Mahaffy et al. 2015). La mejor estima de abundancia para las aguas de Hawái, se basa en los transectos lineares realizados durante 2017, donde se estimaron 12.607 (CV=0,18) individuos (Brandford et al., 2017).
La mejor estima de abundancia para el calderón tropical en aguas del norte del golfo de Méjico es de 1.321 (CV=0,43) individuos (Garrison et al., 2020). Aunque no existen evidencias directas que sugieran la independencia poblacional de la comunidad de calderón tropical en las aguas del Atlántico norte de Estados Unidos, se consideran por separado debido a que se trata de dos ecorregiones marinas diferentes (Spalding et al., 2007). Así, para el calderón tropical en la costa oeste de Estados Unidos (Atlántico norte sin considerar el golfo de Méjico) la mejor estima es de 28.924 (CV=0,24) individuos (Garrison & Palka, 2018).
Los indicios sugieren que también son abundantes en aguas profundas frente a la costa oeste de África, alrededor de las Maldivas y, en general, en el norte del Océano Índico (Minton et al., 2018).
Foto: As. Tonina
Dieta y comportamiento de caza
Es una especie que se alimenta fundamentalmente de cefalópodos, aunque complementa su dieta con diferentes especies de crustáceos y peces (Mintzer et al., 2008; Seagars & Henderson, 1985; Sinclair, 1992) y que muestra diferencias en la dieta dependiendo del área geográfica. Así, en las costas de Carolina del Norte (oeste del Atlántico norte), análisis de contenidos estomacales de esta especie mostraron que la principal presa de cefalópodo era Brachioteuthis riisei (28 % de abundancia numérica), seguida de Taonius pavo (12%) e Histioteuthis reversa (9%), encontrándose asimismo que el pez oceánico Scopelogadus beanii (25 %) era también un componente significativo de la dieta (Mintzer et al., 2008). En el Pacífico, los análisis de contenidos estomacales de calderones varados en California revelaron que se alimentaban fundamentalmente de Loligo opalescens (95 % del total de restos encontrados) (Seagars & Henderson, 1985; Sinclair, 1992), mientras que en japón parece alimentarse de calamares de la Familia Ommastrephidae (potas) y pulpos (Kubodera & Miyazaki, 1983).
En Canarias, aunque el calderón tropical es una de las especies que más aparece en los varamientos (Carrillo & Tejedor, 2002), apenas existen datos sobre sus contenidos estomacales. Hernández-García & Martín (1994) analizaron los contenidos de dos estómagos de calderones en el archipiélago y encontraron restos de cefalópodos meso-batipelágicos (como Discoteuthis sp., Cranchia scabra), lo que coincide con los resultados obtenidos por Fernández et al. (2009) en otro ejemplar varado en Los Gigantes. En un tercer ejemplar se encontraron los restos de un pez de profundidad (Regalecus glesne) (V. Martín com. pers., revisado en Aguilar de Soto, 2006). En Tenerife, es relativamente frecuente que aparezcan pedazos frescos de calamares gigantes (Architeuctis dux) flotando en las cercanías de grupos de calderones, lo que unido a los datos sobre comportamiento de buceo obtenidos utilizando marcas de adherencia, ha llevado a algunos autores a sugerir que esta especie de calamar forma parte de la dieta de los calderones (Aguilar de Soto, 2006; Aguilar de Soto et al., 2008).
El análisis de los perfiles de ácidos grasos de muestras de grasa subcutánea de calderones tropicales, obtenidos en Canarias mediante biopsias, mostró que no existían diferencias significativas en la dieta entre diferentes años o relacionadas con el sexo (Marrero et al., 2016; García-Pastor et al., 2021; Íñiguez-Santamaría et al., 2021).
Los valores medios en composición de isótopos estables de los calderones de Tenerife son: 12,20±0,54 de δ15N y -15,82±0,28 de δ13C (Marrero et al., 2016; Escánez, 2019). Los modelos de contribución a la dieta de los calderones, utilizando los análisis de presas potenciales del calderón y realizados mediante SIAR, revelaron que el calderón tropical en Tenerife parece ser un depredador generalista, no centrando su alimentación sobre ninguna especie en particular (Marrero et al., 2016; Escánez, 2019).
Foto: As. Tonina
Los calderones de Tenerife se alimentan tanto durante el día como por la noche, adaptando el tipo de buceo al tipo de presas y a los ritmos circadianos. Así, durante el día, los calderones realizan buceos profundos (prof. media de 800 m) donde se realizan pocos zumbidos (relacionados con intentos de captura de presas) y éstos se concentran en la parte más profunda de las inmersiones. Por el contrario, durante la noche el calderón realiza intentos de captura en todos los rangos de profundidad (media 300 m), realizando más zumbidos e intentando capturar un mayor número de presas en cada buceo (Aguilar de Soto, 2006; Aguilar de Soto et al., 2008). La combinación sprint-zumbido de los buceos profundos diurnos del calderón define una estrategia de alimentación “tipo guepardo”, que se caracteriza por su elevado coste energético, ya que los animales alcanzan picos de velocidad vertical de 9 m/s (Aguilar de Soto et al., 2008). Los sprints son eventos puntuales de ejercicio intenso, comparables al tipo de esfuerzos que requieren el uso de metabolismo anaeróbico en animales terrestres. Es por ello que se cree que los calderones realizan estas inmersiones diurnas a gran profundidad para cazar cefalópodos de consistencia musculosa, con un alto valor calórico, como pueden ser Todarodes saggitatus o Lepidoteuthis Grimaldi (Aguilar de Soto et al. 2008), que aparecen en sus contenidos estomacales y que alcanzan tamaños de 75 y 97 cm respectivamente (Jereb & Roper, 2010).
Foto: As. Tonina
Reproducción y crecimiento
El calderón tropical, al igual que otras especies de odontocetos, es una especie longeva (~60 años), con madurez tardía, diferentes tasas de maduración para machos y hembras, con apareamiento estacional y el parto de una sola cría en intervalos de varios años (revisado en Bernard & Reilly, 1999).
Las hembras alcanzan la madurez sexual entre los 7-12 años, mientras que los machos la alcanzan a los 13-16 años (Katsuya & Marsh, 1984), aunque éstos últimos parece que logran aparearse con éxito varios años después de alcanzar la madurez. Las diferencias en la tasa de crecimiento hacen que los machos que acaban de alcanzar la madurez sexual sean hasta un 14% más grandes que hembras adultas perfectamente desarrolladas, pudiendo llegar a pesar 2 toneladas y doblando a las hembras en peso (algunas fuentes postulan que los machos pueden llegar a los 3.600 kg.; Jefferson et al., 1993). Los machos son ligeramente más grandes que las hembras al nacer y parecen tener un desarrollo acelerado durante la pubertad (Sergeant, 1962).
El periodo de gestación dura entre 12-15 meses, unos 3 años de lactancia y un periodo entre partos de unos 7 años (Kasuya & Marsh, 1984). La edad media del destete es de unos 4 años, aunque algunas crías permanecen mamando hasta los 6 años, con un máximo registrado de 15 años en un macho inmaduro (Kasuya & Marsh, 1984). Una proporción importante de las hembras de calderón tropical sobreviven más allá de su vida reproductiva, siendo junto con los humanos y otras especies de odontocetos, una de las pocas especies en la que se conoce que pasa por la menopausia (Ellis et al., 2018).
Las hembras parecen vivir más que los machos, 63 y 45 años respectivamente (Kasuya & Marsh, 1984).
Foto: As. Tonina
Comportamiento acústico
Los calderones tropicales producen una gran variedad de sonidos diferentes: chasquidos (“clicks”) y zumbidos (“buzzes”) de eco-localización (Aguilar de Soto et al., 2008) y otros sonidos pulsados y tonales con potenciales funciones comunicativas (Jensen et al., 2011; Pérez et al., 2017).
Ecolocalización
El comportamiento de ecolocalización del calderón tropical es similar al descrito para el cachalote, y los zifios de Cuvier y de Blainville (Miller et al., 2004; Johnson et al., 2004; Zimmer et al., 2005).
Los calderones realizan un proceso de búsqueda y discriminación del alimento en la que emiten largas series de chasquidos, seguida de una fase de aproximación a la presa y, finalmente, un intento de captura, caracterizado por la emisión de un zumbido (Aguilar de Soto, 2006; Aguilar de Soto et al., 2008).
Los zumbidos de los calderones, denominados en inglés “buzz” en delfines y zifios (Au, 1993; Johnson et al., 2004; Aguilar de Soto et al., 2008), y “creak” en la bibliografía de cachalotes (Miller et al., 2004; Watwood et al., 2006), son series de chasquidos emitidos en ráfagas, con un intervalo entre chasquidos (“ICI”) medio de 0.02 s (Aguilar de Soto, 2006), lo que permite a los animales obtener una imagen acústica con más resolución en los momentos previos a la captura de las presas (Johnson et al., 2004; Madsen et al., 2013). Se ha observado que los calderones reducen el ICI de las series regulares de chasquidos progresivamente al aumentar la profundidad durante los buceos, a medida que se acercan a la profundidad a la que emiten un zumbido (que puede utilizarse como estimador de la posición de la presa) (Aguilar de Soto, 2006). Esto parece ser una adaptación a la búsqueda de presas profundas, que están más lejanas al inicio del buceo y se van acercando al calderón, al llegar éste a cotas más profundas (Aguilar de Soto, 2006).
Comunicación
Se han llevado a cabo diferentes estudios acerca del comportamiento acústico de esta especie en el archipiélago canario (P. ej. Scheer, 2013), pero estos trabajos fueron realizados durante cortos encuentros con grupos de calderones en superficie, donde no se describieron apropiadamente las diferentes señales. Trabajos más recientes han utilizado marcas acústicas provistas de múltiples sensores, Dtags (Johnson & Tyack, 2003) para el estudio acústico de los calderones, pudiéndose constatar que esta especie usa señales pulsadas y tonales para comunicarse (Jensen et al., 2011; Sayigh et al., 2012; Marrero, 2015; Pérez et al., 2016).
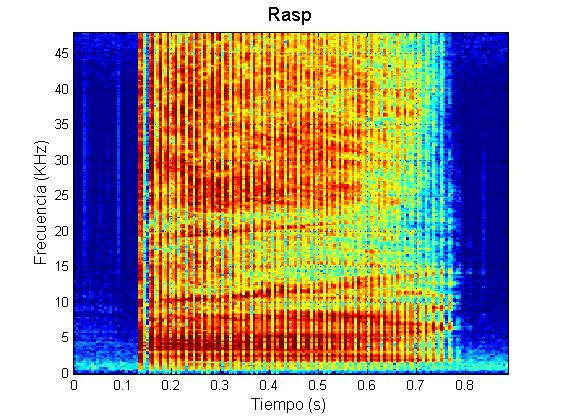
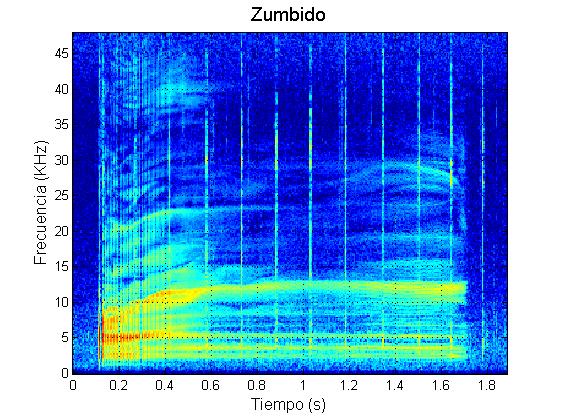
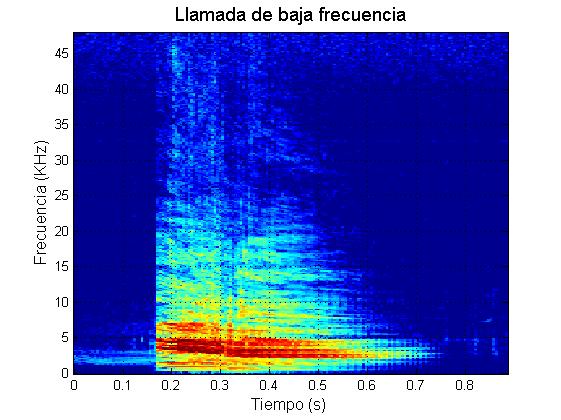
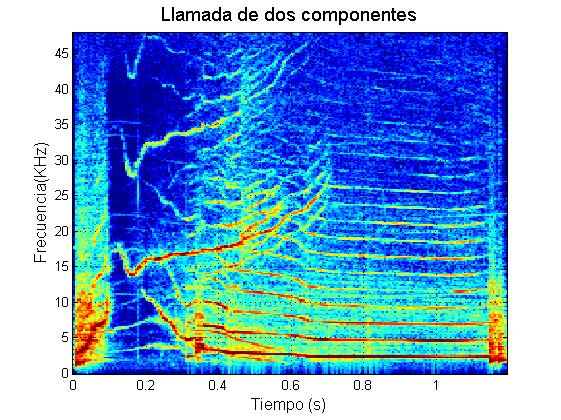
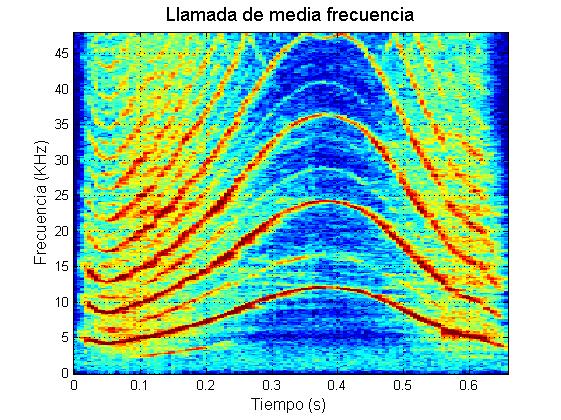
Marrero y colaboradores (2016) caracterizaron el repertorio acústico del calderón tropical en las Islas Canarias. Estos autores descubrieron que durante sus contextos naturales de comportamiento, los calderones tropicales producen cinco tipos principales de vocalizaciones (además de los chasquidos regulares de ecolocalización): zumbidos, rasps, y llamadas de baja frecuencia (“LFC”, de las siglas en inglés: Low Frequency Calls), llamadas de media frecuencia (“MFC”, de las siglas en inglés: Medium Frequency Calls) y llamadas de dos componentes (“TCC”, de las siglas en inglés: Two Component Calls).
Ejemplo rasp
Ejemplo zumbido (buzz)
Los zumbidos se asocian a súbitos incrementos en la tasa de aceleración (“jerk”) del animal que los produce. Estos cambios en la aceleración apoyan la hipótesis de que los zumbidos se asocian a los momentos previos a la captura de presas, de forma homóloga a como sucede en los murciélagos (Johnson et al., 2004; Aguilar de Soto et al., 2008, 2012). Los rasps, aunque son estructuralmente similares a los zumbidos, son más cortos y sus chasquidos se producen a tasas más bajas de repetición que las de los zumbidos. Además, los rasps no se asocian a grandes cambios en la aceleración de los calderones, lo que sugiere que los rasps no están relacionados con procesos de ecolocalización en los momentos previos a la captura de presas (Pérez et al., 2016).
Las llamadas de esta especie fueron clasificadas atendiendo al valor umbral observado en la media de la frecuencia fundamental o de la primera “sideband”, que separa entre los dos tipos de llamadas monofónicas (LFC y MFC, Fig. 3). Inspecciones detalladas de las formas de onda y reproduciendo las grabaciones a 1/10 de su velocidad normal, revelaron que la mayoría de las llamadas de baja frecuencia parecen estar constituidas por pulsos emitidos a alta repetición (Pérez et al., 2016).
Ejemplo llamada de baja frecuencia
Ejemplo llamada de dos componentes
Ejemplo llamada de media frecuencia
Un alto porcentaje de las llamadas de media frecuencia (46%) muestran gradaciones desde trenes de pulsos emitidos extremadamente rápido (frecuentemente al principio de la vocalización) hasta señales sinusoidales continuas de tipo tonal. Resultados similares ya habían sido observados previamente por Sayigh et al. (2012) en calderones tropicales de Bahamas. Esta gradación desde pulsos a tonos continuos parece confirmar la hipótesis de que la frecuencia fundamental de estas llamadas está formada por la vibración neumática de los labios de mono, como ha sido demostrado recientemente para los silbidos del delfín mular (Madsen et al., 2012). Esto contradice la hipótesis de Lilly (1962), que sostenía que la frecuencia fundamental de las señales tonales se genera por la resonancia de los sacos nasales. Un único mecanismo de producción acústica para los calderones tropicales explicaría la aparente continuidad que se observa entre chasquidos regulares, ráfagas de pulsos (zumbidos y rasps) y llamadas; fenómeno observado también en otros delfínidos (Murray et al., 1998).
Las llamadas de dos componentes de los calderones tropicales poseen dos frecuencias fundamentales (con sus armónicos asociados) superpuestas en el tiempo, lo que sugiere que estos sonidos son producidos por la vibración simultánea de los dos pares de labios de mono, como parece ocurrir en otras especies de odontocetos como la orca (Tyson et al., 2007) de modo que cada uno de los componentes sería emitido por uno de los pares de labios fónicos.
Debido a que la tasa de producción de las llamadas aumenta durante la fase de ascenso de los buceos profundos del calderón tropical, se cree que este tipo de señales juegan papeles importantes para el restablecimiento del contacto entre los individuos que están buceando y los que permanecen en superficie (Jensen et al., 2011; Marrero, 2015; Pérez et al., 2016).
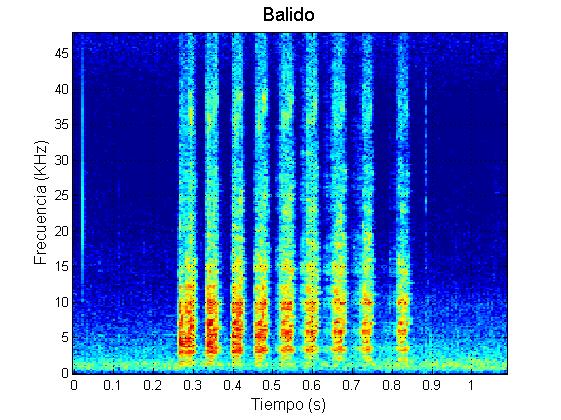
Además, los calderones tropicales de Tenerife producen en ocasiones otros tipos de vocalizaciones. Algunas, como el llamado “balido“, no han sido descritas aún en la bibliografía y se han encontrado en grabaciones de machos.
Ejemplo balido
Estructura social
Utilizando la definición dada por Bigg y coautores (1990) para definir los grupos sociales de orcas, Heimlich-Boran (1993) agrupó los calderones fotoidentificados en Tenerife en grupos familiares básicos. Este autor observó un total de 46 grupos, con un tamaño medio de 12,2 individuos en cada grupo. De esos grupos, 31 fueron considerados residentes, con una media de 14 ind./grupo, y 15 grupos fueron considerados visitantes, con un tamaño medio de 8 ind./grupo. Además, estos grupos se clasificaron como grupos productivos y no productivos, en base a si presentaban crías o juveniles en los mismos. Se observó que los grupos productivos presentaban más animales por grupo (15,5 ind./grupo) frente a los no productivos (4,6 ind./grupo) (Heimlich-Boran, 1993).
El modelo de “compañeros constantes”, que indica un sistema social con relaciones duraderas entre sus miembros, parece ser el que mejor predice los patrones de asociación del calderón tropical en los trabajos realizados en Hawaii y Madeira (Mahaffy, 2012; Alves et al., 2013). Alves y colaboradores (2013) analizaron datos genéticos paralelamente a los datos de fotoID, obteniendo que los miembros de los distintos grupos sociales están estrechamente emparentados entre sí, lo que parece indicar que existe una alta filópatria natal en esta especie. Este estudio parece confirmar la hipótesis de Heimlich-Boran (1993) de que los grupos sociales de calderón tropical tienen un tipo de organización social jerarquizada de tipo matrilineal, donde los machos no se reproducen con las hembras de sus mismos grupos sociales, con quienes están emparentadas por vía materna, para evitar la endogamia (Heimlich-Boran, 1993; Alves et al., 2013). En el caso de la comunidad de calderón tropical del suroeste de Tenerife se observan grupos numerosos durante primavera-verano (Heimlich-Boram, 1993; Montero & Martín, 1993; Carrillo & Tejedor, 2002). Estos grandes grupos parecen estar formados por asociaciones temporales de dos o más unidades familiares y se cree que se forman para favorecer el apareamiento (Heimlich-Boram, 1993; Alves et al., 2013).
Los datos de fotoID de los calderones más frecuentemente avistados en Tenerife y La Gomera mostraron también un sistema social con asociaciones duraderas entre sus miembros (Servidio, 2014), donde se identificaron 11 grupos que permanecían estables a lo largo del tiempo, pero en los que se encontraban niveles de asociación variables, lo que sugiere que existe cierto grado de intercambio entre los miembros de algunos de los grupos (Servidio, 2014).
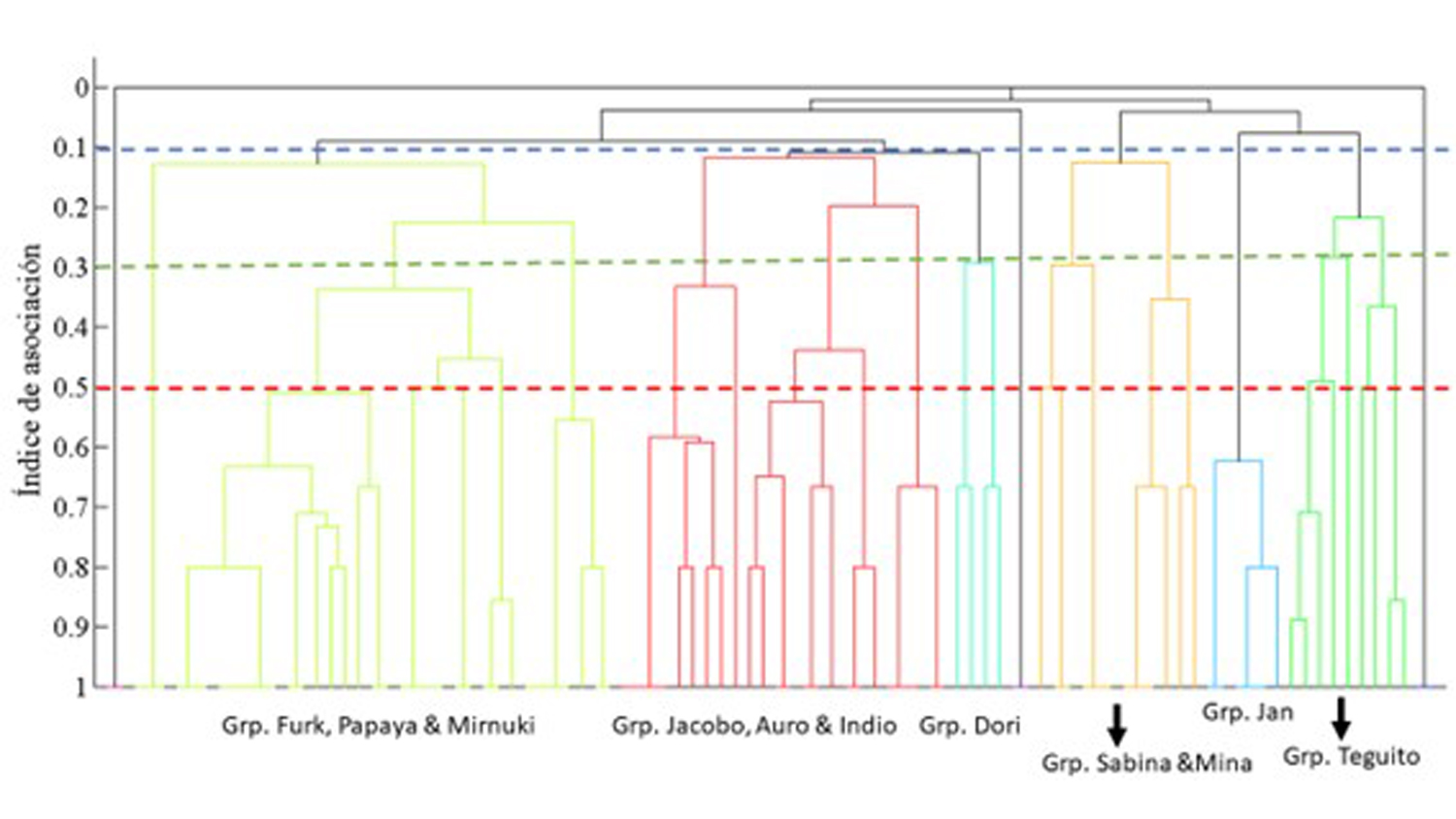
Recientes análisis de la estructura social de los calderones “residentes” del suroeste de Tenerife evidenciaron las dificultades que existen para definir correctamente lo que es un “grupo social” en esta especie, donde se observa de forma habitual la dispersión y posterior unificación de individuos por la zona (Pimentel, 2020). Se demuestra en cualquier caso la existencia de relaciones no aleatorias entre individuos, siendo el modelo de conocidos casuales el que mejor se ajusta a los datos, con 6 (2-22 individuos) ó 9 (2-17 individuos) grupos sociales identificados, dependiendo del criterio utilizado para definir los grupos durante la toma de datos (Pimentel, 2020).
Figura 4. Cluster de grupos sociales presentes en los calderones residentes del Sw de Tenerife.
El calderón tropical en Canarias
En las aguas del archipiélago canario se pueden producir avistamientos de calderón tropical en todas las islas, siendo relevante la presencia de una comunidad residente establecida en aguas costeras del suroeste de Tenerife (Heimlich-Boran, 1993; Montero & Martín, 1993; Carrillo & Tejedor, 2002; Servidio, 2014; Marrero et al., 2016; Marrero & Fernández, 2018; Servidio et al., 2019). Esta comunidad no se encuentra aislada del resto de la población canaria de calderones, existiendo flujo de animales de unas islas a otras (Servidio, 2014; Servidio et al., 2019). Además, los reavistamientos de individuos observados entre los archipiélagos de Madeira y Azores (5-8 % de la población), y entre Canarias y Madeira (0,2 % de la población), confirman que existe un flujo de individuos entre los archipiélagos de la Macaronesia (Alves et al., 2019).
Durante los muestreos archipelágicos realizados por La Universidad de La Laguna (2009-2010) se observó una alta detección visual de grupos de calderón tropical (n=31) en las aguas colindantes a la isla de Tenerife, sobre todo en el suroeste y este de la isla. En otras zonas del archipiélago, a pesar de las buenas condiciones de mar y visibilidad durante los muestreos, sólo se registraron avistamientos aislados (Fais et al., 2010).
Los últimos muestreos archipelágicos realizados por CEAMAR durante 2017, obtuvieron asimismo una alta proporción de avistamientos de esta especie (n=49) alrededor de la isla de Tenerife, aunque también se observaron grupos de calderón tropical entre Gran Canaria y Fuerteventura, y al sur-suroeste de La Gomera (MISTIC SEAS II, 2019). Utilizando la metodología “Distance sampling” se realizaron estimas de 135 animales (95% CI=38-277) para Gran Canaria, 545 (95% CI=173-699) para La Gomera, 1.082 (95% CI=642-1.456) para Tenerife y 577 (95% CI=228-1.146) para el canal entre Gran Canaria y Fuerteventura (MISTIC SEAS II, 2019). Con una densidad de 0,022 individuos por km2 para Gran Canaria, de 0,151 indv. por km2 para La Gomera, de 0,137 indv. por km2 para Tenerife y de 0,176 por km2 para el canal entre Gran Canaria y Fuerteventura (MISTIC SEAS II, 2019).
Foto: As. Tonina
La población residente del suroeste de Tenerife
La comunidad de calderón tropical establecida en el suroeste de Tenerife se considera residente, ya que pueden producirse avistamientos durante todo el año y algunos de los grupos sociales que se avistan regularmente mantienen una alta fidelidad por un área catalogada como Zona de Especial Conservación: Franja Marina Teno-Rasca (ZEC ES-7020017) protegida por la directiva europea Hábitat (Heimlich-Boran, 1993; Montero & Martín, 1993; Carrillo & Tejedor, 2002; Servidio, 2014; Marrero et al., 2016; Marrero & Fernández, 2018; Servidio et al., 2019). Esta comunidad es de gran importancia a nivel mundial, ya que sólo se conocen otras poblaciones residentes en: Japón, California, Hawai, Madeira y las Marianas (Heimlich-Boran, 1993; Kasuya & Tai, 1993; Olson, 2009; Mahaffy, 2012; Alves et al., 2013; Mahaffy et al, 2015; Hill et al., 2019).
En el año 1989, Heimlich-Boran realizó el primer estudio de los calderones en esta zona, dándola a conocer para la ciencia. Durante los 22 meses que duró este estudio, se registró una presencia diaria de calderones tropicales de 0,77 avistamientos por unidad de esfuerzo al día (APUE/día) (Heimlinch-Boran, 1993). Estudios posteriores llevados a cabo de 1992 a 1993 y de 2001 a 2002 obtuvieron resultados de 0,92 avistamientos/día (Montero & Martín, 1993) y 3,57 avistamientos/día (Carrillo & Tejedor, 2002) respectivamente. Esta diferencia en los APUE/día puede explicarse, al menos en parte, debido a las diferentes embarcaciones empleadas en los distintos estudios, habiéndose empleado embarcaciones neumáticas hasta el estudio llevado a cabo por Carrillo y Tejedor, donde se empleó un barco de 13 m de eslora con una plataforma de avistamiento situada a 7 m de altura.
Ninguno de los estudios realizados hasta la fecha han demostrado la existencia de variaciones estacionales significativas en la frecuencia de avistamientos de esta especie (Heimlich-Boran, 1993; Montero & Martín, 1993; Carrillo & Tejedor, 2002; Servidio, 2014). No obstante, parecen existir incrementos en los APUE/día a lo largo del año, que parecen variar entre los diversos estudios llevados a cabo (realizados en años diferentes). Así, se detectaron ligeros incrementos en las APUE/día de abril a junio (Montero, 1997) y de diciembre a enero (Carrillo & Tejedor, 2002). Este último resultado coincide con los valores obtenidos por Morales (2015), en el que se utilizó un modelo ARIMA para el análisis de series temporales de avistamientos y donde se observaron incrementos en el número de días con avistamientos de calderones tropicales durante el otoño.
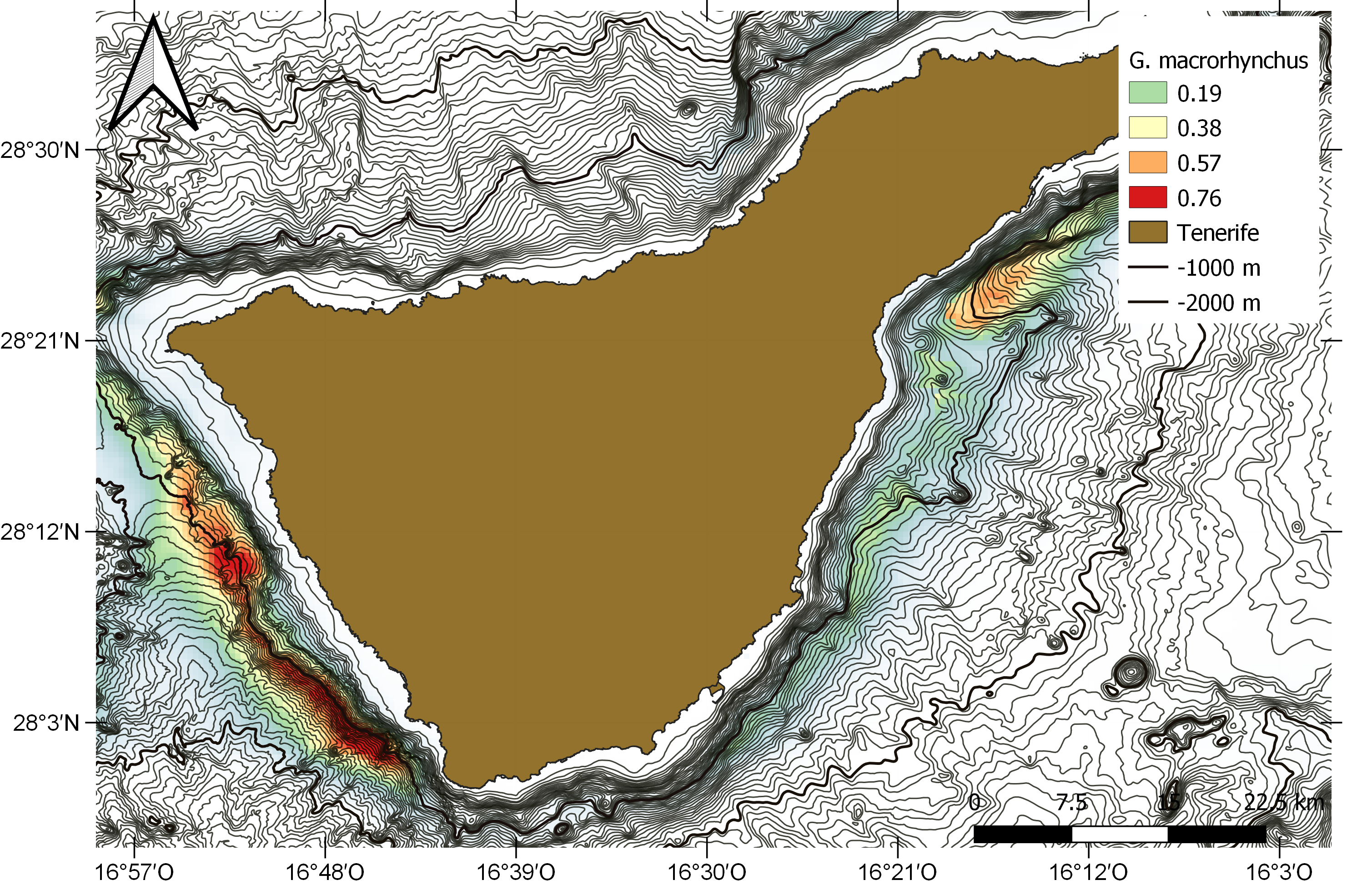
La distribución de los avistamientos de calderón tropical en el área del ZEC (S-W de Tenerife) no es homogénea, existiendo zonas de mayor o menor densidad en base a la frecuencia de avistamientos en el área (Fig. 2). Todos los estudios realizados coinciden en situar una preferencia de los grupos de calderones en torno a la isóbata de 1.000 m (Heimlich-Boran, 1993; Montero & Martín, 1993; Carrillo & Tejedor, 2002; Servidio, 2014; Marrero et al., 2016) (Fig. 2).
Figura 2. Modelo de nicho ecológico de calderón tropical (Globicephala macrorhynchus) en Tenerife.
Utilizando técnicas de fotoidentificación, Heimlich-Boran (1993) elaboró un catálogo con 495 ejemplares (más 77 animales inmaduros sin marcas características claras). Utilizando la frecuencia de individuos reavistados, los animales catalogados fueron clasificados en 388 residentes (individuos que aparecían más de una vez) y 107 visitantes (que sólo fueron avistados una vez). Un estudio posterior, utilizando la misma metodología, obtuvo un número similar de calderones residentes, aunque 117 fueron clasificados como visitantes (Montero & Martín, 1993). Hay que tener en cuenta que éstos fueron estudios acumulativos de individuos durante el periodo de estudio (1989 a 1993) y no pueden considerarse en ningún caso como estimas del número de calderones en el área.
Una estima de abundancia en la que se utilizó la metodología de “Distance sampling”, obtuvo un valor medio de 362 individuos (CI=241-544) para el área, con una densidad de 1,47 individuos por Km2 (Carrillo & Tejedor, 2002).
Utilizando datos obtenidos desde 1999 a 2012, Servidio y colaboradores clasificaron a 50 calderones fotoidentificados en Tenerife como residentes usuales y a 246 como residentes, de un total de 711 individuos (Servidio et al., 2019). Estos autores descubrieron variaciones estacionales en el tamaño de los grupos en la zona, siendo los grupos mayores en otoño y primavera (Servidio et al., 2019).
Las últimas estimas realizadas (años 2015, 2017, 2018, 2019 y 2020) utilizando técnicas de captura-recaptura por fotoidentificación, muestran que la comunidad de calderones tropicales del suroeste de Tenerife se ha mantenido relativamente estable durante todos estos años, con 362 indv. residentes (ICI=252-746) (Marrero et al., 2016; Marrero & Fernández, 2018; Pimentel et al., 2021; García-Pastor et al., 2021).
Foto: As. Tonina
Bibliografía
-Aguilar de Soto, N. (2006). Acoustic and foraging behavior of short-finned pilot whales (Globicephala macrorhynchus) and Blainville’s beaked whales (Mesoplodon densirostris) in the Canary Islands. Implications for impacts of man-made noise and ship collisions. Phd. Thesis. La Laguna University, Canary Islands, Spain.
-Barlow, J. (2016). Cetacean abundance in the California current estimated from ship-based line-transect surveys in 1991-2014. Southwest Fisheries Science Center, Administrative Report, LJ-2016-01. 63 p.
-Bernard, H. J. & Reilly, S. B. (1999). Pilot whales Globicephala Lesson, 1828 in: Handbook of Marine Mammals. Volume 6: The Second Book of Dolphins and Porpoises. Academic Press, San Diego.
-Bloch, D., Zachariassen, M. & Zachariassen, P. (1993). Some external characters of the long-finned pilot whale off Faroe Island and a comparison with the short-finned pilot whale. Rep. Int. Whal. Comm.(Special Issue) 14, 117-135.
-Bradford, A.L., K.A. Forney, E.M. Oleson, and J. Barlow. (2017). Abundance estimates of cetaceans from a line-transect survey within the U.S Hawaiian Islands Exclusive Economic Zone. Fishery Bulletin 115:129142.
-Cañadas, A. (2006). Towards conservation of dolphins in the Alborán Sea. Phd thesis, Universidad Autónoma de Madrid.
-Carrillo, M. & Tejedor, M. (2002). Estudios aplicados a la conservación de cetáceos en la provincia de Santa Cruz de Tenerife. Gobierno de Canarias. Consejería de política territorial y Medio Ambiente. Viceconsejería de Medio Ambiente.
-Forney, K. A. (1997). Patterns of variability and environmental models of relative abundance for California cetaceans. Ph.D. Dissertation, Scripps Institution of Oceanography, University of California, San Diego.
-Fraser, F. C. (1950). Two skulls of Globicephala macrorhyncha (Gray) from Dakar. Atlantide Report. 1, 49-60.
-García Pastor, E., Crespo Torres, A., Pimentel, A., Montañés, A., Iñiguez Santamaría, E., Escánez Pérez, A., García Oliva B. M. y Marrero Pérez, J. (2021). Cambios en la población residente de calderón tropical (Globicephala macrorhynchus) del suroeste de Tenerife asociados al cese de actividad náutica de recreo durante el confinamiento decretado durante la crisis sanitaria Covid-19. Memoria técnica de Asociación Tonina para Tragsatec por encargo de la Dirección General de Lucha Contra el Cabio Climático y Medio Ambiente de la Consejería de Transición Ecológica, Lucha Contra el Cambio Climático y Planificación Territorial del Gobierno de Canarias.
-Jones Jr, J. K., Carter, D. C., Genoways, H. H., Hoffmann, R. S., Rice, D. W., & Jones, C. (1986). Revised checklist of North American mammals north of Mexico. Occ Papers Mus Texas Tech Univ 107:5.
-Kasuya, T. & Marsh, H. (1984). Life history and reproductive biology of the short-finned pilot whale, Globicephala macrorhynchus, off the Pacific Coast of Japan. Rep. Int. Whal. Comm. 6, 239-310.
-Kasuya, T. & Tai, S. (1993). Life history of short-finned pilot whale stocks of Japan and a description of the fishery. In Biology of Northern Hemisphere Pilot Whales. Report of the International Whaling Commission, pp. 439-473. International Whaling Commission, Cambridge.
-Kubodera, T., & Miyazaki, N. (1993). Cephalopods eaten by short‐finned pilot whales, Globicephala macrorhynchus, caught off Ayukawa, Ojika Peninsula, in Japan, in 1982 and 1983. In Recent advances in cephalopod fisheries biology (pp. 215-227). Tokai University Press Tokyo.
-Marrero Pérez, J. & Fernández, M. (2018). Estudios para la realización de un censo de la población de calderón tropical (Globicephala macrorhynchus) en la ZEC Franja Marina Teno-Rasca (Tenerife). Informe de Asociación Tonina para el Ministerio de Agricultura, Alimentación y Medio Ambiente.
-Miyashita, T., Kishiro, T., Higashi, N., Sato, F., Mori, K. & Kato, H. (1996). Winter distribution of cetaceans in the western North Pacific inferred from sighting cruises 1993-1995. Report of the International Whaling Commission 46, 437-442.
-Montañés Pérez, A., García Pastor, E., Pimentel, A. y Marrero Pérez, J. (2021). Estudio del ruido subacuático generado por las embarcaciones que realizan la AROC en la ZEC Teno-Rasca y gradación de las características más óptimas de éstas. Memoria técnica de GAIA consultores para el Ministerio para la Transición Ecológica y el Reto Demográfico.
-Montero López, R. & Martín Martel, V. (1993). Estudio del impacto que provocan las embarcaciones en la población de calderones tropicales (Globicephala macrorhynchus) residente en aguas del suroeste de la isla de Tenerife, pp. 136, Santa Cruz de Tenerife.
-Montero, R. (1997). Actividades de observación de mamíferos marinos. Consejería de transporte del Gobierno de Canarias y Sociedad para la promoción del Turismo, Naturaleza y Ocio.
-Morales, T. (2015). Análisis de la distribución estacional de cetáceos en las Islas Canarias. Master thesis, Universidad de La Laguna.
-Pimentel, A. (2020). Estima poblacional y estructura social de calderón tropical (Globicephala macrorhynchus) del suroeste de Tenerife, Islas Canarias. Master thesis, Universidad de La Laguna.
-Pimentel, A., Montañés, A., García Pastor, E. y Marrero Pérez, J. (2021). Determinación de los niveles de cortisol en 10 ejemplares de calderón tropical de la ZEC Franja Marina Teno-Rasca. (Memoria técnica). Informe de Asociación Tonina para el Ministerio para la Transición Ecológica y el Reto Demográfico.
-Red de Varamientos de cetáceos en Canarias, 2000-2019. Base de datos del Gobierno de Canarias. Origen de los datos: ULPGC-IUSA (Universidad de Las Palmas de Gran Canaria-Instituto Universitario de Sanidad Animal). Islas Occidentales: Manuel Carrillo, Tenerife Conservación-GEA (Gabinete de Estudios ambientales). Islas Orientales: Marisa Tejedor, SECAC (Sociedad para el Estudio de los Cetáceos en el Archipiélago Canario).
-Van Bree, P. J. H. (1971). On Globicephala sieboldii Gray, 1846, and other species of pilot whales (Notes on Cetacea, Delphinoidea III). Beaufortia 19, 79-87.